

Seguro que muchos de los que habéis leído el título de este post pensaréis que os voy a dar una chapa infernal sobre plantas y Botánica, e incluso algunos/as tendréis la idea de que las plantas son los seres vivos más aburridos de todo el planeta, porque no hacen nada más que estar tiesas y realizar la fotosíntesis (idea equivocada).
Hoy vengo a desmontar este mito de que las plantas son lo más soso que existe, y os demostraré que, al contrario de lo que pueda parecer en un principio, las plantas son organismos increíbles, con una capacidad de adaptarse al medio y una resistencia envidiables.
¿Qué son las plantas exactamente?
Las plantas se pueden definir como aquellos seres vivos multicelulares cuyas células deben poseer una pared celular rígida y cloroplastos que les permitan realizar la fotosíntesis (1).
Obviamente, estas características no son exclusivas de las plantas, ya que existen levaduras que poseen pared celular, y también hay algas que poseen cloroplastos y pared celular (2).
Ahora, muchos os estaréis preguntando: ¿y por qué las algas no son plantas, si las hay multicelulares, con cloroplastos y pared celular? Pues básicamente no se consideran plantas por varias razones: primero, no están tan diferenciadas como las plantas, es decir, que carecen de algunas estructuras vegetales como son las flores, raíces, tallos... (3). Además, los organismos clasificados como algas son muy diversos entre sí, ya que podemos encontrar algas unicelulares (cosa que no ocurre con las plantas) y multicelulares, a lo que se suma el hecho de que las algas ocupan mayoritariamente hábitats acuáticos (al contrario que las plantas, que se encuentran principalmente en ecosistemas terrestres).
De hecho, hay organismos procariotas, como las cianobacterias, que pueden ser clasificados dentro del grupo de las algas y son capaces de realizar la fotosíntesis, y además cabe destacar que, según la teoría endosimbiótica de Lynn Margulis, los cloroplastos de las células eucariotas vegetales aparecieron gracias al establecimiento de una relación de endosimbiosis entre células eucariotas primitivas y cianobacterias, que a lo largo de la evolución fueron dando lugar a los cloroplastos actuales (4). Es decir, que las plantas actuales surgieron a partir de algas.

Aquí tenéis a las cianobacterias, a partir de las cuales surgieron los cloroplastos en las células eucariotas vegetales, gracias a la relación de endosimbiosis que establecieron con los ancestros de las células eucariotas actuales (Fuente).
Las plantas se pueden clasificar en dos tipos principales: las que viven en ecosistemas acuáticos, como es el caso de las fanerógamas marinas -o las ya mencionadas algas, en caso de considerarlas plantas- y las plantas terrestres o embriófitas, que se pueden clasificar a su vez en plantas vasculares o traqueófitas (es decir, que poseen tejidos vasculares como el xilema y floema) y en briófitas (que no poseen tejidos vasculares, como los musgos) (5) (6). Hoy nos centraremos en las plantas vasculares o traqueófitas, las más "avanzadas" evolutivamente, y ya con esto terminamos con la parte más tediosa del post (¡bieeeen!).
La resistencia de las plantas al estrés
Como ya comenté al principio del post, una de las cosas que hacen a las plantas tan chulas es su capacidad de adaptarse de forma asombrosa a las adversidades del medio y a los cambios que se producen en el mismo, pudiendo así sobrevivir en condiciones que en ocasiones llegan a ser extremas. Esto es así porque, como bien sabéis, las plantas no son capaces de desplazarse de su sitio, y por ello deben poseer una serie de mecanismos y estrategias que les permitan sobrevivir y adaptarse ante cualquier cambio en las condiciones ambientales o ante los distintos tipos de estrés a los que se ven sometidas. Además, todo esto viene acompañado de una complejidad fisiológica y metabólica alucinantes que hacen que las plantas puedan responder de forma adecuada ante una gran variedad de situaciones adversas.
Para hacernos una idea clara de este concepto, el estrés hace referencia a cualquier condición medioambiental adversa que interfiera en el normal desarrollo y crecimiento de la planta (7).
En concreto, el estrés que puede sufrir una planta se puede clasificar en dos tipos principales: estrés biótico (aquel debido al ataque de patógenos como hongos, bacterias, virus o herbívoros) y estrés abiótico (el que se debe a factores como la sequía, salinidad, radiaciones, metales pesados o temperaturas extremas) (8).
Vamos a centrarnos en este post en el estrés abiótico, ya que supone la principal causa de pérdidas en los cultivos a nivel global, y por ello tiene un enorme impacto tanto a nivel económico como social.

Nota importante: cuando hablamos de estrés biológico -en este caso, en plantas-, NO ME REFIERO A ESTE TIPO DE ESTRÉS, ¿VALE? ¿DE ACUERDO? (Fuente).
El estrés abiótico: tipos principales
En primer lugar, vamos a hablar de cómo afecta la sequía a las plantas: el agua, como todos sabréis, es esencial para que se pueda desarrollar la vida y es fundamental que su disponibilidad sea la óptima para los seres vivos, y las plantas no son una excepción en este aspecto.
Para hablar del estrés hídrico y la disponibilidad del agua, hemos de tener claro un concepto de química física denominado potencial hídrico, que nos va a ayudar a comprender un poco mejor los mecanismos y procesos que ocurren a nivel molecular durante los periodos de sequía, y cómo las plantas pueden hacer frente a ellos.
Antes de definir el potencial hídrico, hemos de comprender qué es el potencial químico (µ): este concepto hace referencia a la energía libre por mol que contiene una sustancia química, o entendiéndolo de otra manera (un poco más compleja de comprender), se puede definir como el incremento de energía libre que experimenta un sistema químico al añadir una cantidad determinada de esa especie química (9). El potencial químico es un indicador de la energía potencial que posee una sustancia en solución, y depende de una serie de factores, como pueden ser la composición química, la entropía (que mal dicho, se puede entender como el "desorden" del sistema), la presión, la temperatura y el potencial eléctrico (en el caso de que la especie química posea carga) (10).
El potencial hídrico (φ) se puede definir básicamente como el potencial químico que posee el agua en un medio o sistema con unas condiciones determinadas (11), y su valor hace referencia a la energía potencial que posee el agua en una solución determinada con respecto al agua pura (sin solutos ni otras sustancias, y en unas condiciones estándar).
El potencial hídrico que posee el agua pura en condiciones estándar tiene por convenio un valor de 0, que además supone el máximo valor posible que puede alcanzar el potencial hídrico, con lo cual, cualquier sistema acuoso que posea solutos u otras sustancias químicas poseerá un valor de φ negativo. Es importante saber el potencial hídrico que poseen dos compartimentos que estén comunicados, ya que se producirá de forma espontánea (sin aporte de energía externa) un flujo de agua desde el compartimento que posea un valor mayor de φ hasta el que tenga menor valor de φ.

Fórmula del potencial hídrico (φ): vemos en esta expresión cómo el potencial hídrico de una solución acuosa depende de diversas variables, como el potencial osmótico (debido a la presencia de solutos), el potencial mátrico (que depende de las características del suelo o matriz en la que se encuentre el agua), la presión hidrostática y el potencial gravitatorio (que aumentará a mayor altitud) (Fuente).
Imagina que tenemos dos compartimentos: A y B, cuyos potenciales hídricos son φA= -0.6 MPa y φB= -0.9 MPa. La diferencia de potencial hídrico entre B y A es de 0.3 MPa, siendo menor el valor de φ en el compartimento B, por lo que en el caso de que el agua pueda fluir de un lado hacia otro (a través, por ejemplo, de una membrana que deje pasar el agua), se producirá un flujo de agua neto desde A hasta B. Además, cuanto mayor sea la diferencia de φ entre dos compartimentos, mayor será la tendencia que tendrá el agua a pasar del sitio con mayor φ al sitio de menor φ. Un ejemplo de esto lo podemos encontrar, por ejemplo, cuando se produce el flujo de agua desde el suelo hacia las células de la raíz de una planta.
En el caso de que tengamos un suelo con abundante humedad, este puede poseer un valor de φS = -0.01 MPa (12), mientras que el potencial hídrico de una célula de la raíz puede rondar un valor de φR = -0.2 MPa. Como vemos, el valor del potencial hídrico de una célula radicular es bastante menor (más negativo) que el del suelo, así que se producirá de forma espontánea un flujo neto de agua desde el suelo hacia las raíces de la planta.
Además, en este principio de diferencias de potencial hídrico se basa también el transporte de agua desde el tallo de la planta hasta las células de las hojas, que poseen un menor valor de φ que las células del tallo. Así que como veis, es de gran importancia que las células que forman la planta posean unos valores de φ determinados para que se dé el transporte de agua y sustancias de una parte a otra de la planta.

Potencial hídrico en diferentes partes de la planta. Como veis, el potencial hídrico va disminuyendo a medida que vamos ascendiendo desde las raíces de la planta hasta sus hojas. Esto permite -junto a procesos de capilaridad- que el agua llegue a todas las partes de la planta a pesar de la fuerza gravitatoria, que dificulta el ascenso del agua a través del tallo. Además, vemos que el valor de φ de la atmósfera es mucho más bajo que en la planta, de ahí que se den de forma espontánea los procesos de evaporación de agua y transpiración. (Fuente).
Lo que ocurre en situaciones de sequía, es que la concentración de agua del suelo es menor que en condiciones normales, por lo que el valor de φS disminuye de forma notable, haciendo que la absorción de agua a través de la raíz sea menor lo cual supone un problema para nuestras pobres plantas.
Este tipo de estrés, conocido también como estrés hídrico, no solo puede deberse a la falta de precipitaciones, sino a temperaturas extremas. Cuando hace mucho calor, la planta está continuamente perdiendo agua por procesos de evaporación y transpiración (la salida de vapor de agua a través de unas "aperturitas" que tienen las hojas, llamadas estomas), y además, el potencial hídrico de los suelos disminuye al haber menor concentración de agua en ellos.
Sin embargo, también el frío causa estrés hídrico en las plantas, aunque nos parezca algo "raro" a priori: esto es debido a que, a muy bajas temperaturas, pueden aparecer cristales de hielo en el medio extracelular, lo que hace que ese agua solidificada no esté "disponible" al estar formando hielo, y esto hace que disminuya de forma notable el valor de φ del medio extracelular, lo que provoca finalmente un flujo neto de agua desde las células hacia el medio externo.
Además, las temperaturas extremas (frío o calor) no solo provocan estrés hídrico en las plantas, sino que alteran el funcionamiento de las mismas de otras maneras.
Las temperaturas muy bajas pueden afectar a las plantas no solo a través de la disminución de la disponibilidad de agua al formar cristales de hielo, ya que esos mismos cristales de hielo pueden dañar seriamente la estructura de las células vegetales, como pueden ser las membranas celulares, provocando una ruptura de las mismas a medida que esos cristales de hielo se forman y van creciendo. Esto, a su vez, causa la "fuga" de iones de la célula a través de las zonas donde ha habido rupturas de la membrana plasmática, lo que compromete seriamente muchísimas funciones celulares ya que la membrana plasmática, además de regular los gradientes iónicos entre las células y el exterior, también desempeñan funciones de señalización y regulación del tráfico de nutrientes y metabolitos (13). Además, el frío hace que pueda aumentar o disminuir la tasa metabólica de la planta, se producen metabolitos atípicos y esto conlleva, entre otras cosas, un crecimiento o maduración del fruto anormales, aparición de manchas en los frutos e incluso la muerte de la planta.
En el otro extremo, tenemos aquellas condiciones de calor extremo, en las que aparte del estrés hídrico, se producen otra serie de alteraciones que alteran las funciones de las células vegetales. Por ejemplo, las altas temperaturas se cree que favorecen un crecimiento más rápido de las plantas, alteran su desarrollo y pueden influenciar en el proceso de germinación de las semillas.

Las temperaturas extremas pueden provocar estrés hídrico, pero también afectan a diferentes niveles a las plantas, alterando su fisiología y su metabolismo (Fuente).
Este tipo de estrés por calor, junto al estrés hídrico asociado al mismo, no son problemas leves ni mucho menos, pues hay cultivos que pueden perderse en más de un 50% debido a condiciones de sequía y calor, lo cual supone pérdidas económicas muy severas para los productores e incluso pueden dar lugar a problemas de hambre en regiones empobrecidas (14).
Además del estrés térmico (por frío o calor), también se puede producir estrés por salinidad. Este tipo de estrés se debe a un aumento excesivo de la concentración de sales en el suelo donde se encuentran los vegetales.
Podemos clasificar el estrés por salinidad en dos tipos: primario, cuando este se debe a causas naturales, como una época de bajas precipitaciones, o secundario, cuando se debe a acciones humanas, como ocurre cuando se utilizan fertilizantes de forma abusiva o cuando se utilizan aguas de riego salinizadas.
Los principales efectos debidos a una alta salinidad del medio son dos: la toxicidad iónica y el estrés osmótico.
La toxicidad iónica hace referencia a aquellas situaciones en las que la concentración de ciertos minerales en los suelos -como Na+, K+ o Ca2+- alcanza valores tan elevados que impiden el normal crecimiento o desarrollo de los cultivos (15).
El estrés osmótico ocurre normalmente en condiciones de sequía o altas temperaturas, lo que produce un aumento en la concentración de sales en el suelo, aumentando de esta forma su presión osmótica. Cuando el aumento de la salinidad del medio llega a niveles considerables, podemos llegar a un punto en el que la presión osmótica del suelo sea más elevada que la de las células de las raíces, haciendo entonces que el potencial hídrico sea menor en el suelo que en la planta, y entonces se dé un flujo neto de agua desde la planta al suelo (ya que el potencial hídrico depende de la presión osmótica). Esto hace que la planta no pueda absorber agua del suelo, lo que junto a los procesos de pérdida de agua -principalmente por transpiración y evaporación- provoca que la planta vea detenido su crecimiento y su metabolismo se vea alterado.

Representación esquemática del proceso de osmosis. La osmosis tiene lugar cuando dos soluciones con diferentes concentraciones de solutos -o presiones osmóticas- están en contacto a través de una membrana semipermeable, que permite el paso de disolvente (agua), pero no de los solutos. Se produce así un flujo neto de agua desde el compartimento que posee mayor presión osmótica (esto es, mayor concentración de solutos y menor valor de φ) hasta el compartimento con menor presión osmótica. El flujo de agua se detiene una vez se han igualado las presiones osmóticas (concentraciones de solutos) de ambas soluciones (Fuente).
Los principales efectos del estrés salino se manifiestan a muchos niveles: por ejemplo, la germinación de las semillas en plantas como el trigo se ve bastante afectada en condiciones de estrés salino (16), lo cual supone un problema debido a la gran importancia que tiene esta especie a nivel agroalimentario. De hecho, se piensa que este estrés salino que puede sufrir el trigo provoca la acumulación de unos compuestos llamados ROS, o especies reactivas de oxígeno, que son muy dañinos para las células ya que poseen una gran avidez por los electrones presentes en otras moléculas -como el DNA o las proteínas celulares-, pudiendo arrebatárselos y dañando así muchas de las estructuras celulares (17).
Ya por último, comentaremos brevemente en qué consiste el estrés por metales pesados.
Dentro del grupo de los metales pesados encontramos aquellos elementos de la tabla periódica que son metales y poseen una masa atómica mayor o igual a 20. Normalmente, estos metales pesados se encuentran de forma natural en los suelos, y algunos de ellos, como el manganeso (Mn), cobre (Cu) o zinc (Zn), son micronutrientes (véase el post anterior) y ejercen funciones en las células vegetales, mientras que otros como el mercurio (Hg) o el plomo (Pb) no tienen una función biológica conocida (18).
La acumulación de alguno de estos metales pesados en las plantas (o en cualquier ser vivo) es tóxica, y puede acarrear consecuencias negativas ya que pueden alcanzar niveles peligrosos en los seres vivos o en los propios ecosistemas al no ser biodegradables (19).

Un pequeño inciso: una de las mayores preocupaciones de los consumidores de pescado es la contaminación con metales pesados. La acumulación de metales pesados (sobre todo de mercurio, Hg) supone un problema para la industria pesquera y para los consumidores de pescado, sobre todo de aquellas especies de mayor tamaño como el atún, el tiburón o el pez espada, cuyos niveles de Hg son mayores ya que se alimentan de otros peces de menor tamaño que ya poseen mercurio en sus organismos (Fuente).
¿Cómo hacen las plantas para defenderse ante el estrés abiótico?
En esta sección vamos a centrarnos únicamente en cinco mecanismos de defensa que utilizan la mayor parte de las plantas ante los distintos tipos de estreses abióticos que pueden sufrir a lo largo de su ciclo vital: la cutícula, los ácidos grasos insaturados (UFAs), la eliminación o scavenging de ROS, la activación de chaperonas moleculares y la producción de solutos compatibles (20).
1. Cutícula:
La cutícula es una capa cerosa formada por una matriz de cutina en la cual se encuentran embebidas una serie de ceras cuticulares, y que se encuentra sobre la epidermis de las plantas ejerciendo una función de protección de las mismas. Seguro que alguna vez habréis visto fotos en las que se observan gotitas de agua muy "perfectas" y redonditas sobre las hojas de una planta, ¿verdad? Pues la formación de esas gotas de agua que se deslizan sobre la superficie de las hojas sin llegar a mojarlas es posible gracias a la presencia de la cutícula sobre las células epidérmicas de las hojas.

Repulsión del agua por parte de la cutícula. Estas imágenes tan chulas se pueden observar en las hojas de las plantas gracias a la presencia de la cutícula. Al ser de naturaleza hidrofóbica, impide que las moléculas de agua "mojen" las hojas de las plantas (Fuente).
Obviamente, las funciones biológicas de la cutícula poco tienen que ver con fines estéticos (la naturaleza no existe ni mucho menos para hacernos disfrutar a los humanos), sino que su papel es esencial en la protección de la planta frente a la deshidratación y otros tipos de estrés biótico (ataque de patógenos) o abiótico (compuestos químicos, radiación UV...) (21).
En cuanto a su composición química, la cutina -el principal constituyente de la cutícula- consiste en un polímero (poliéster) de ácidos grasos oxigenados de 16 o 18 átomos de carbono, mientras que las ceras cuticulares son mezclas de derivados de ácidos grasos de entre 24 y 34 átomos de carbono, entre los que se encuentran cetonas, aldehídos, alcoholes, ésteres o alcanos.
La síntesis de novo de estos compuestos que forman la cutícula y que son básicamente moléculas derivadas de los ácidos grasos se inicia en los cloroplastos, donde se produce la síntesis de los ácidos grasos en las células vegetales. Posteriormente, los ácidos grasos sintetizados se transportan al retículo endoplasmático, donde sufren una serie de modificaciones en función de si se utilizarán para sintetizar la matriz de cutina o las ceras que se encuentran dentro de ella. Una vez sintetizados los derivados de ácidos grasos que formarán la cutícula, estos son transportados a través de la membrana plasmática y la pared celular gracias a unas proteínas transportadoras de lípidos, que depositan los componentes de la cutícula en el medio extracelular (donde finalmente polimerizarán dando lugar a la cutina y las ceras de la cutícula, gracias a una serie de reacciones enzimáticas y no enzimáticas).
Los mecanismos por los cuales se da la polimerización de la cutina no están aún claros del todo: de hecho, se ha visto que plantas mutantes conocidas como cd1 (cutin-defficient 1), que poseen inactivado el gen que codifica la proteína responsable de sintetizar la cutina, son aún así capaces de fabricar cutina, con lo cual se cree que hay otros genes sin descubrir de la misma familia que también intervienen en la síntesis de cutina, o que pueden existir mecanismos no enzimáticos adicionales que intervienen en la síntesis de este polímero (22).
2. Ácidos grasos insaturados (UFAs):
Los ácidos grasos insaturados o UFAs, por sus siglas en inglés (unsaturated fatty acids), no solo intervienen en la defensa de la planta ante el estrés al ser precursores de los componentes de la cutícula epidérmica, sino que también actúan a nivel de las membranas celulares (sobre todo, en la membrana plasmática que delimita el citoplasma del medio extracelular).
En concreto, los UFAs son esenciales a la hora de regular la fluidez de las membranas, porque al aumentar su concentración dentro de las mismas harán que las membranas celulares sean más fluidas, ya que los UFAs -al poseer dobles enlaces en cis que provocan una ligera curvatura de la cadena hidrocarbonada- presentan un menor grado de empaquetamiento, y por tanto, interacciones intermoleculares más débiles que hacen que las membranas sean más inestables y su fluidez sea mayor.
Esto tiene una gran relevancia en la supervivencia de las células cuando estas se encuentran en condiciones de temperaturas extremas, ya que a muy bajas temperaturas, las membranas celulares presentan menor fluidez debido a la disminución de la energía térmica, mientras que a altas temperaturas, la fluidez aumenta notablemente, con lo cual es necesario modular la proporción de ácidos grasos saturados e insaturados en las membranas en función de las condiciones de temperatura en las que se encuentren las plantas (lo que estas deben evitar es que sus membranas cristalicen a bajas temperaturas, y que se desestabilicen a altas temperaturas).
Además, a esto se añade el hecho de que la gran mayoría de las plantas son organismos poiquilotermos, es decir, que al contrario que nosotros los seres humanos (organismos homeotermos), su temperatura corporal varía con la del ambiente (23), de ahí que sea de enorme importancia regular estrictamente la fluidez de las membranas celulares en función de la temperatura ambiental.
Pero no solo eso, ya que a partir de los ácidos grasos, las plantas pueden sintetizar una serie de compuestos bioactivos de gran relevancia en la respuesta ante el estrés, conocidos como oxilipinas. Entre ellas, destaca un grupo de moléculas con función hormonal y señalizadora llamadas jasmonatos, todos ellos derivados del ácido jasmónico (JA).

Yo no sé vosotros, pero a mí la palabra "jasmónico" me hace pensar automáticamente en esto (Fuente).
El JA es esencial en la regulación del crecimiento y la respuesta ante el estrés de las plantas, junto a otra serie de hormonas, como el etileno, las auxinas, las giberelinas o los brassinosteroides (24). Por ejemplo, cuando una planta sufre el ataque de un insecto o una herida, se incrementan los niveles endógenos de JA, activando así una vía de señalización que culmina en la activación de unos factores de transcripción (TFs) de la familia MYC, que regulan la expresión de genes que están implicados en la respuesta ante situaciones de estrés (por ejemplo, mediante la apertura o cierre de estomas) (25).
3. Eliminación o scavenging de especies reactivas de oxígeno (ROS):
Las especies reactivas de oxígeno (ROS) son una serie de moléculas y radicales que poseen una alta reactividad y poder oxidante, ya que necesitan electrones para alcanzar una mayor estabilidad. Por ello, las ROS se dedican a ir por ahí robando electrones a otras moléculas para alcanzar ellas su ansiada estabilidad (unas manguis, vamos). Esto, como estaréis imaginando, supone un serio problema cuando las ROS están presentes en alta cantidad en una célula, pues esas ROS podrán robar electrones a las moléculas de agua del medio o a otras biomoléculas -como el DNA, las proteínas o los lípidos-, dañando así componentes celulares que posean importantes funciones en la célula.
Las ROS, a pesar de lo chungas que puedan parecer, se encuentran presentes en muy pequeñas concentraciones dentro de las células de una gran cantidad de seres vivos en un estado fisiológico, debido a la actividad respiratoria mitocondrial, o en vegetales debido también a la cadena de transporte electrónico tilacoidal. Además, al estar habitualmente a muy bajas concentraciones, las ROS ejercen una función de señalización muy importante en las células, regulando así una serie de procesos entre los cuales se encuentran algunos mecanismos de resistencia ante el estrés. Lo que sucede es que en situaciones adversas, se empiezan a acumular altas cantidades de ROS que pueden suponer un peligro para las células, con lo cual se activan una serie de metabolitos y enzimas conocidas como ROS scavengers (o "eliminadores" de ROS) que se encargan de regular los niveles de especies reactivas de oxígeno con los fines de regular diferentes rutas de señalización o de evitar lesiones en el DNA o daños en otras biomoléculas. De hecho, las ROS pueden activar vías de señalización que modulan la expresión de genes implicados en infinidad de procesos como la senescencia, la diferenciación celular, el desarrollo o el crecimiento (26).
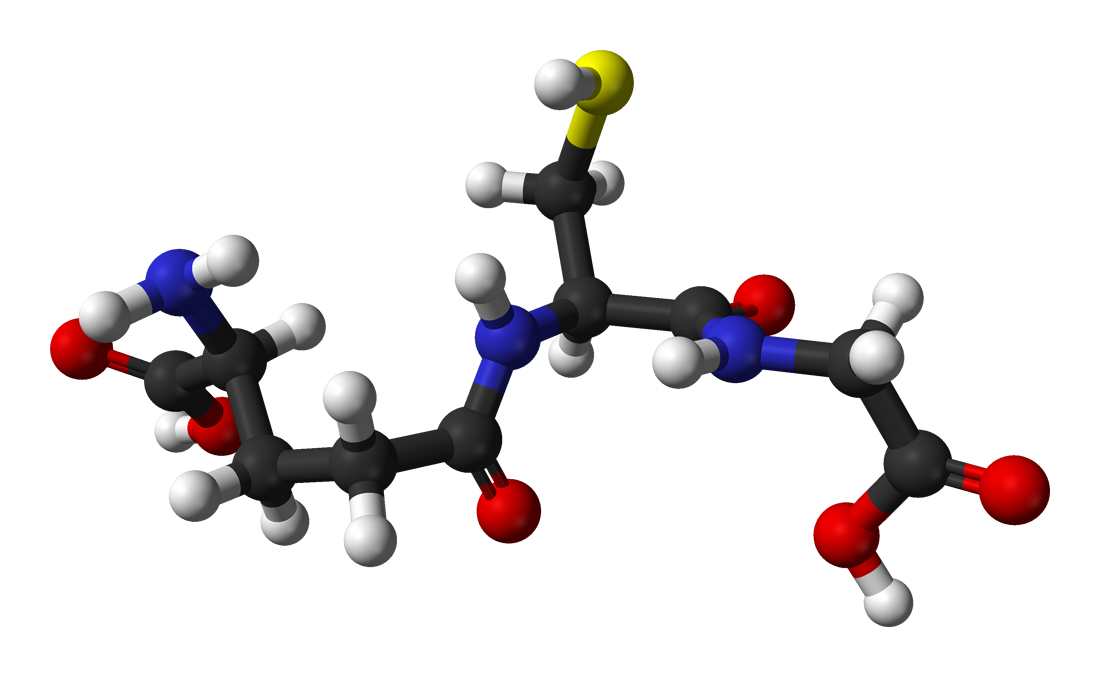
Os presento a uno de los scavengers de ROS por excelencia, el glutation. Este tripéptido de glutamato, cisteína y glicina es uno de los principales antioxidantes celulares, el cual mencionaremos posteriormente (Fuente).
4. Chaperonas moleculares:
Las chaperonas son una serie de proteínas importantísimas en las células, ya que asisten el proceso de plegamiento de otras muchas proteínas, y las mantienen plegadas -o desplegadas- cuando es necesario. Una curiosidad que aprendí este año en clase es que la palabra chaperona viene de América Latina, donde allí se conocían como chaperones o chaperonas a aquellas personas "que acompañan a una pareja o a una joven (olé ahí ese machismo bueno) para vigilar su comportamiento", según dice la RAE.
Se puede hacer la analogía de las chaperonas "de humanos" con las de proteínas, ya que en este caso se encargan de controlar que las proteínas no se queden mal plegadas una vez sintetizadas o que mantengan el estado de plegamiento indicado en cada momento.
De hecho, es tal la importancia de las chaperonas que se encuentran prácticamente en todos los organismos vivos. En el caso que nos atañe -nuestras queridas plantas- hay un tipo de chaperonas llamadas HSPs (heat-shock proteins), entre las que destaca HSP70, la chaperona más conservada entre los diferentes seres vivos. Estas proteínas se encargan, como ya hemos visto, de asistir el plegamiento de las proteínas sintetizadas, o ayudar a que proteínas mal plegadas o parcialmente plegadas adquieran un correcto plegamiento. Además, HSP70 se encarga de degradar proteínas mal plegadas (que normalmente presentan residuos de aminoácidos hidrofóbicos expuestos al medio) y que tienden a formar agregados entre ellas, evitando así el daño que supondría la precipitación y acumulación de esos agregados insolubles. Para unirse y separarse de las proteínas que deben ser plegadas o degradadas, la chaperona HSP70 debe hidrolizar y unir moléculas de ATP, gastando energía en el proceso. Para que esto se lleve a cabo, HSP70 requiere además de la "ayuda" de otras co-chaperonas, como HSP40, que posee dominios que estimulan la actividad ATPasa (de hidrólisis de ATP) de HSP70, y también se requieren de factores de intercambio de nucleótido (NEFs) para que HSP70 pueda unirse a moléculas de ATP, y una vez estas sean hidrolizadas gracias a la actividad ATPasa estimulada por HSP40, se pueda liberar el ADP resultante con el fin de poder volver a unir ATP.
También existen otro tipo de chaperonas conocidas como smHSP, cuya diversidad es mayor que la de HSP70 y pueden actuar en diferentes orgánulos de la planta y reconocer también diferentes tipos de proteínas a plegar o degradar. Además, estas smHSP aumentan drásticamente su expresión en situaciones de estrés, llegando a unirse a proteínas recién traducidas para marcarlas y que no sean degradadas por HSP70, sino que sean renaturalizadas por esta chaperona con el fin de asegurar un correcto plegamiento proteico en situaciones adversas, donde es habitual que se formen agregados de proteínas mal plegadas cuya presencia compromete la supervivencia celular (27).

Mecanismo de acción conjunto de las chaperonas HSP70 y HSP40 durante el plegado de una proteína. El proceso requiere de un gasto de energía -en forma de ATP- y de la acción de un factor de intercambio de nucleótido (NEF), pudiendo así plegarse correctamente o no la proteína diana (en este último caso, puede repetirse el ciclo para tratar de plegarla bien, o bien marcarse para su degradación a través del proteosoma) (Fuente).
5. Producción de solutos compatibles:
Los solutos compatibles son compuestos orgánicos que pueden encontrarse a altas concentraciones dentro de las células sin que ello suponga un problema para las mismas. Algunos de los requisitos que debe cumplir una molécula para ser un soluto compatible son: baja toxicidad, electroneutralidad, bajo peso molecular y una elevada solubilidad.
En situaciones de estrés, estos compuestos suelen acumularse y ejercen un efecto osmoprotector ante la deshidratación, actúan como scavengers de ROS, y estabilizan proteínas y membranas lipídicas. De hecho, se ha visto que mutantes de los genes que codifican enzimas implicadas en la degradación de solutos compatibles poseen una mayor resistencia a diferentes tipos de estrés abiótico como el frío, la sequía y la salinidad (28) (29); incluso se han documentado mutantes de Arabidopsis thaliana que son capaces de resistir a temperaturas de bajo cero y ambientes salinos al mismo tiempo (30).
Hay diversos ejemplos de solutos compatibles, como algunos azúcares (rabinosa, trehalosa, rafinosa) y aminoácidos o sus derivados (prolina o glicina-betaína).

El aminoácido prolina puede actuar como metabolito osmoprotector, aumentando así su concentración en situaciones de estrés hídrico (Fuente).
Las hormonas vegetales
No quiero acabar este post sin comentaros la relevancia que poseen las distintas hormonas vegetales o fitohormonas en la adaptación de las plantas a las condiciones ambientales.
Como mencionamos antes, hay hormonas -como el ácido jasmónico o JA- que regulan a nivel molecular el funcionamiento de las células vegetales para que estas se adapten a los distintos tipos de estreses.
Aparte del ácido jasmónico y sus derivados, también existen otras hormonas que juegan un papel fundamental en la respuesta de las plantas ante los diversos tipos de estrés: el ácido abscísico (ABA), el etileno (ET), el ácido salicílico (SA), las auxinas, las giberelinas (GAs), los brassinosteroides y los propios jasmonatos (JAs) (31).
1. Ácido abscísico (ABA): se trata de la principal hormona implicada en la respuesta ante el estrés, ya que interacciona con muchas otras fitohormonas y moléculas señalizadoras, y además, promueven la movilización de moléculas implicadas en la defensa ante estreses como son los precursores de las ceras cuticulares, las chaperonas y los solutos compatibles. También regulan la expresión de una gran cantidad de genes implicados en la respuesta a los distintos tipos de estrés.
El ácido abscísico es sintetizado a partir del ꞵ-caroteno, a través de una ruta metabólica que tiene lugar en los cloroplastos y el citoplasma (32), en la que el paso limitante es la reacción catalizada por la enzima 9-cis-epoxicarotenoidedioxigenasa (o mejor, NCED para los amigos).
2. Etileno (ET): el etileno es una hormona gaseosa implicada principalmente en el control del crecimiento y el desarrollo de las plantas, controlando procesos como la senescencia de las flores o la maduración del fruto, actuando en conjunto con el ABA en muchas ocasiones. Además, a altas concentraciones promueve la tolerancia de las plantas ante el estrés.
Seguro que habréis oído hablar del etileno: cuando dejáis en el frutero -sobre todo en verano- aquellas frutas que producen altas cantidades de esta fitohormona, como pueden ser los plátanos, al poco tiempo veis cómo el resto de la fruta que tenéis junto a los plátanos han madurado considerablemente, llegando en ocasiones a ponerse "malas" y todo (con el coraje que da eso...). Pues el responsable de que se produzca este fenómeno es el etileno, que promueve la maduración de los frutos.

El etileno es una de las fitohormonas responsables de la maduración del fruto. Si dejamos un plátano maduro junto a otras frutas, al ser el etileno un compuesto volátil, es capaz de acelerar la maduración del resto de piezas del frutero (Fuente).
3. Ácido salicílico (SA): otra molécula que seguro que os sonará, ya que todos hemos recurrido alguna vez a ella para aliviar algún dolor de cabeza o para reducir la fiebre cuando nos hemos tomado la típica Aspirina®, cuyo principio activo es este compuesto producido por árboles como el sauce (de ahí su nombre).
Aunque siendo estrictos, lo que realmente constituye el principio activo de la aspirina es un derivado del SA llamado ácido acetilsalicílico, que actúa como inhibidor de unas enzimas llamadas ciclooxigenasas (33) responsables de producir a partir del ácido araquidónico unos compuestos proinflamatorios llamados prostaglandinas, causantes de síntomas como la fiebre o el dolor.
En las plantas, la función del ácido salicílico está relacionada con las respuestas ante el estrés frente a patógenos, es decir, ante el estrés biótico. Sin embargo, no se conoce muy bien su rol en la respuesta ante el estrés abiótico. Básicamente se encarga de regular la expresión de genes implicados en la respuesta ante patógenos, como aquellos que codifican chaperonas o enzimas implicadas en la síntesis de metabolitos secundarios y antioxidantes. A altas concentraciones, puede provocar la muerte celular.

La tan socorrida Aspirina® tiene como principio activo ácido acetilsalicílico, un compuesto derivado de la fitohormona ácido salicílico (SA), cuya función principal en plantas es promover la tolerancia a estreses bióticos (Fuente).
4. Auxinas: estas hormonas ejercen un efecto regulador del crecimiento y desarrollo de la planta, regulando la expresión de genes implicados en ambos procesos, y además, modula las respuestas ante estreses como la presencia de metales pesados o una alta salinidad.
5. Giberelinas (GAs): las giberelinas también regulan los procesos de crecimiento y desarrollo de la planta junto al ácido abscísico, además de promover la tolerancia ante el estrés abiótico como la sequía.
6. Brassinosteroides: los brassinosteroides son, como su propio nombre indica, fitohormonas esteroideas responsables del crecimiento de algunas zonas de la planta como la raíz o el tallo, y promueven la floración y la resistencia ante estreses abióticos como las temperaturas extremas, la sequía, el estrés por luz y los contaminantes. Estas hormonas activan unas vías de señalización que regulan la actividad de diversos factores de transcripción implicados en la expresión de genes relacionados con múltiples procesos, como el crecimiento o el desarrollo.
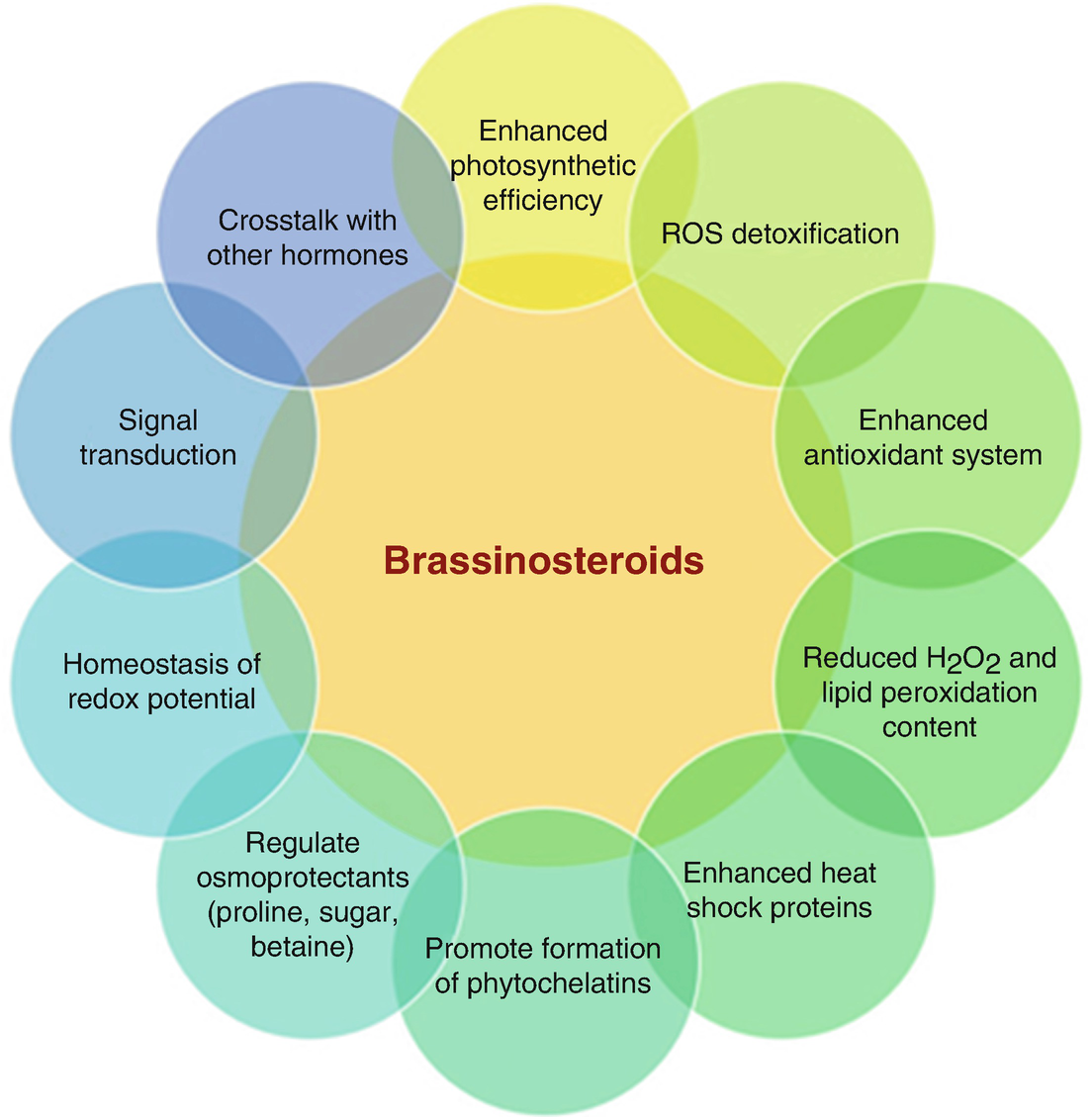
Los brassinosteroides poseen una gran diversidad funcional. Como vemos en este esquema, los brassinosteroides ejercen una gran cantidad de funciones relacionadas con el crecimiento, el desarrollo, y la tolerancia de la planta ante estreses principalmente abióticos (Fuente).
7. Jasmonatos (JAs): por último, vamos a repasar qué funciones llevan a cabo el ácido jasmónico y sus derivados, los jasmonatos.
Como vimos antes, los jasmonatos son derivados oxigenados de los ácidos grasos, que ejercen un efecto regulatorio de la respuesta ante el estrés biótico y abiótico, aunque también son capaces de regular otros procesos fisiológicos como el crecimiento, la formación del fruto y la flor, la senescencia, y el metabolismo secundario.
Cabe destacar el caso del metiljasmonato (MeJA), cuya acumulación tiene lugar en las células de la raíz ante el estrés salino, aumentando así la resistencia de la planta en estas condiciones. Además, el MeJA también reduce el estrés por contaminantes, como son los metales pesados, al inducir la maquinaria celular antioxidante y una serie de péptidos llamados fitoquelatinas (34), constituidos por repeticiones del tripéptido antioxidante glutation y que son capaces de acomplejar metales pesados como el Cu (cobre) o el Cd (cadmio).

El ácido jasmónico (JA) y sus amigos, los jasmonatos. Todos ellos son fitohormonas de gran relevancia en las plantas, ya que intervienen en la tolerancia al estrés, el crecimiento y el desarrollo (Fuente).
Conclusiones
Como habéis podido ver, las plantas son unas auténticas cracks, ya que poseen una capacidad de adaptación asombrosa gracias a su diversidad metabólica y los diferentes mecanismos que poseen para responder adecuadamente a los diferentes tipos de estrés -biótico y abiótico-, pero aún así, no son inmortales, ya que se puede llegar a un punto en el que la fisiología de las plantas colapse y no puedan sobrevivir. Esto supone un serio problema a nivel agroeconómico y agroalimentario, ya que las pérdidas causadas por diferentes tipos de estrés -sobre todo abióticos- son millonarias a nivel mundial, por lo que la investigación en biotecnología, genética y biología molecular vegetal es de vital importancia para que en el futuro podamos dar de comer sin problemas a la creciente población humana y que los agricultores reciban los beneficios que se merecen por el duro trabajo que llevan a cabo.
Espero que os haya gustado mucho este post, y si es así, ¡seguidme en mis cuentas de Instagram (@elbiolocomolecular) y Twitter (@el_bioloco)! ¡Un saludo a todos/as!
Referencias
2. Domozych, D. (2019). Algal Cell Walls. In eLS (pp. 1–11). https://doi.org/10.1002/9780470015902.a0000315.pub4
4. Martin, W. F., Garg, S., & Zimorski, V. (2015). Endosymbiotic theories
for eukaryote origin. In Philosophical Transactions of the Royal Society B:
Biological Sciences (Vol. 370, Issue 1678). https://doi.org/10.1098/rstb.2014.0330
7. Yadav, S.,
Modi, P., Dave, A., Vijapura, A., Patel, D., & Patel, M. (2020). Effect of
Abiotic Stress on Crops. In Sustainable Crop Production. https://doi.org/10.5772/intechopen.88434
8. Gull, A.,
Ahmad Lone, A., & Ul Islam Wani, N. (2019). Biotic and Abiotic Stresses in
Plants. In Abiotic and Biotic Stress in Plants. https://doi.org/10.5772/intechopen.85832
10. Nobel, P. S.
(2009). Chapter 2: Water. In Physicochemical and Environmental Plant
Physiology (4th ed.). Elsevier Academic Press.
13. Takahashi,
D., Li, B., Nakayama, T., Kawamura, Y., & Uemura, M. (2013). Plant plasma
membrane proteomics for improving cold tolerance. In Frontiers in Plant
Science (Vol. 4, Issue APR). https://doi.org/10.3389/fpls.2013.00090
14. Lamaoui, M.,
Jemo, M., Datla, R., & Bekkaoui, F. (2018). Heat and drought stresses in
crops and approaches for their mitigation. Frontiers in Chemistry, 6.
https://doi.org/10.3389/fchem.2018.00026
15. Flowers, T.
J., Munns, R., & Colmer, T. D. (2015). Sodium chloride toxicity and the
cellular basis of salt tolerance in halophytes. In Annals of Botany
(Vol. 115, Issue 3, pp. 419–431). https://doi.org/10.1093/aob/mcu217
16. Gholamali Akbari, Seyed
Ali Mohammad Modarres Sanavy and Saeed Yousefzadeh, 2007. Effect of Auxin and
Salt Stress (NaCl) on Seed Germination of Wheat Cultivars (Triticum
aestivum L.). Pakistan Journal of Biological Sciences, 10: 2557-2561.
17. Wani, S. H.,
Tripathi, P., Zaid, A., Challa, G. S., Kumar, A., Kumar, V., Upadhyay, J.,
Joshi, R., & Bhatt, M. (2018). Transcriptional regulation of osmotic stress
tolerance in wheat (Triticum aestivum L.). In Plant Molecular Biology
(Vol. 97, Issue 6, pp. 469–487). https://doi.org/10.1007/s11103-018-0761-6
18. Tangahu, B.
V., Sheikh Abdullah, S. R., Basri, H., Idris, M., Anuar, N., & Mukhlisin,
M. (2011). A review on heavy metals (As, Pb, and Hg) uptake by plants through
phytoremediation. In International Journal of Chemical Engineering. https://doi.org/10.1155/2011/939161
19. Azimi, A.,
Azari, A., Rezakazemi, M., & Ansarpour, M. (2017). Removal of Heavy Metals
from Industrial Wastewaters: A Review. In ChemBioEng Reviews (Vol. 4,
Issue 1, pp. 37–59). https://doi.org/10.1002/cben.201600010
20. He, M., He,
C. Q., & Ding, N. Z. (2018). Abiotic stresses: General defenses of land
plants and chances for engineering multistress tolerance. In Frontiers in
Plant Science (Vol. 871). https://doi.org/10.3389/fpls.2018.01771
21. Ziv, C.,
Zhao, Z., Gao, Y. G., & Xia, Y. (2018). Multifunctional roles of plant
cuticle during plant-pathogen interactions. In Frontiers in Plant Science
(Vol. 9). https://doi.org/10.3389/fpls.2018.01088
22. Fich, E. A.,
Segerson, N. A., & Rose, J. K. C. (2016). The Plant Polyester Cutin:
Biosynthesis, Structure, and Biological Roles. In Annual Review of Plant Biology
(Vol. 67, pp. 207–233). https://doi.org/10.1146/annurev-arplant-043015-111929
24. Yang, J.,
Duan, G., Li, C., Liu, L., Han, G., Zhang, Y., & Wang, C. (2019). The
Crosstalks Between Jasmonic Acid and Other Plant Hormone Signaling Highlight
the Involvement of Jasmonic Acid as a Core Component in Plant Response to
Biotic and Abiotic Stresses. In Frontiers in Plant Science (Vol. 10). https://doi.org/10.3389/fpls.2019.01349
25. Gimenez-Ibanez,
S., Boter, M., Ortigosa, A., García-Casado, G., Chini, A., Lewsey, M. G.,
Ecker, J. R., Ntoukakis, V., & Solano, R. (2017). JAZ2 controls stomata
dynamics during bacterial invasion. New Phytologist, 213(3). https://doi.org/10.1111/nph.14354
26. Huang, H.,
Ullah, F., Zhou, D. X., Yi, M., & Zhao, Y. (2019). Mechanisms of ROS
regulation of plant development and stress responses. In Frontiers in Plant
Science (Vol. 10). https://doi.org/10.3389/fpls.2019.00800
27. Bao, Y.,
& Howell, S. H. (2017). The unfolded protein response supports plant
development and defense as well as responses to abiotic stress. Frontiers in
Plant Science, 8. https://doi.org/10.3389/fpls.2017.00344
28. Sun, Z., Qi,
X., Wang, Z., Li, P., Wu, C., Zhang, H., & Zhao, Y. (2013). Overexpression
of TsGOLS2, a galactinol synthase, in Arabidopsis thaliana enhances tolerance
to high salinity and osmotic stresses. Plant Physiology and Biochemistry,
69, 82–89. https://doi.org/10.1016/j.plaphy.2013.04.009
29. Zhuo, C.,
Wang, T., Lu, S., Zhao, Y., Li, X., & Guo, Z. (2013). A cold responsive
galactinol synthase gene from Medicago falcata (MfGolS1) is induced by myo-inositol
and confers multiple tolerances to abiotic stresses. Physiologia Plantarum,
149(1). https://doi.org/10.1111/ppl.12019
30. Nanjo, T.,
Kobayashi, M., Yoshiba, Y., Kakubari, Y., Yamaguchi-Shinozaki, K., &
Shinozaki, K. (1999). Antisense suppression of proline degradation improves
tolerance to freezing and salinity in Arabidopsis thaliana. FEBS Letters,
461(3). https://doi.org/10.1016/S0014-5793(99)01451-9
31. El‐Esawi, M.
A. (2017). Introductory Chapter: Hormonal Regulation in Plant Development and
Stress Tolerance. In Phytohormones - Signaling Mechanisms and Crosstalk in
Plant Development and Stress Responses. https://doi.org/10.5772/intechopen.69806
32. Cardoso, A.
A., Gori, A., Da-Silva, C. J., & Brunetti, C. (2020). Abscisic acid
biosynthesis and signaling in plants: Key targets to improve water use
efficiency and drought tolerance. In Applied Sciences (Switzerland)
(Vol. 10, Issue 18). https://doi.org/10.3390/APP10186322




{kind=link}
{kind=link}
{kind=link}